

This article explores the characteristics, causes, and unknowns of pattern hair loss – also known as androgenic alopecia (AGA).
If you’re new to hair loss research, this is a good starting point to learn the science behind AGA as well as the concepts that come up frequently in the literature.
- Hair follicle miniaturization
- Telogen:anagen ratios
- Dihydrotestosterone, 5-alpha reductase, androgen receptors, and androgen receptor co-activation
- Prostaglandin activity
- Wnt-ß catenin pathways
- Future research focuses: olfactory receptors, galea interaction, and more
After all, the better we understand a condition, the better equipped we are to treat it.
We’ll explore how the interplay between genetics and male hormones may lead to hair follicle miniaturization. Then, we’ll reveal what still puzzles researchers about AGA, despite 50+ years of study.
Finally, we’ll dive into more recent evidence on AGA that is paving a new direction of research – and reshaping the future of treatments.
What is androgenic alopecia?
Androgenic alopecia (AGA), also known as pattern hair loss, is one of the most common hair loss disorder in the world. It’s chronic, progressive, and affects up to 50% of women and 80% of men at some point in their lives. In the U.S. alone, 30 million women and 50 million men are dealing with AGA that is severe enough to become diagnosable (NIH).
What does AGA look like?
AGA can occur both in men and women, but it manifests differently between the two sexes.
In men, classic pattern balding appears as:
- Recession of the hairline at the temples
- Loss of hair at the vertex (crown)
Norwood-Hamilton Scale (Male Pattern Hair Loss)
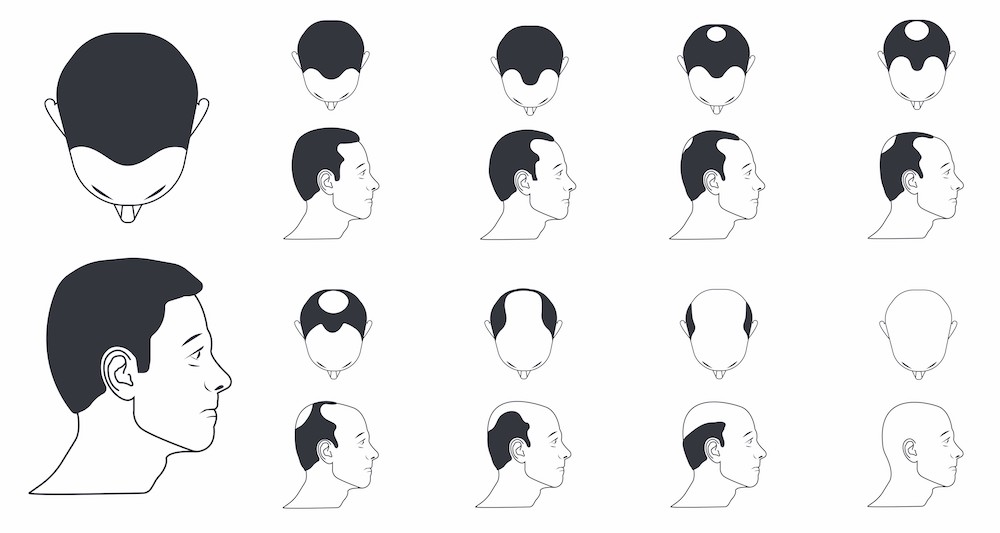

However, female pattern hair loss can present in a few different ways (Herskovitz et al., 2013):
- Diffuse, even thinning of the crown with preservation of the front hairline
- Christmas tree-pattern hair loss, with a widening of the scalp beginning at the front and tapering towards the crown
- Thinning at the temples
Ludwig Scale (Female Pattern Hair Loss)


What are the hallmark characteristics of AGA?
AGA has specific features that distinguish it from other types of hair loss.
For starters, it primarily affects hair at the top part of the scalp – above the galea aponeurotica (the dense fibrous membrane that stretches across the top of the scalp).
Secondly, scalp regions affected by AGA show three defining characteristics of the condition:
- Hair follicle miniaturization
- Increased telogen:anagen hairs
- Shortened anagen cycling
Each of these phases are covered below.
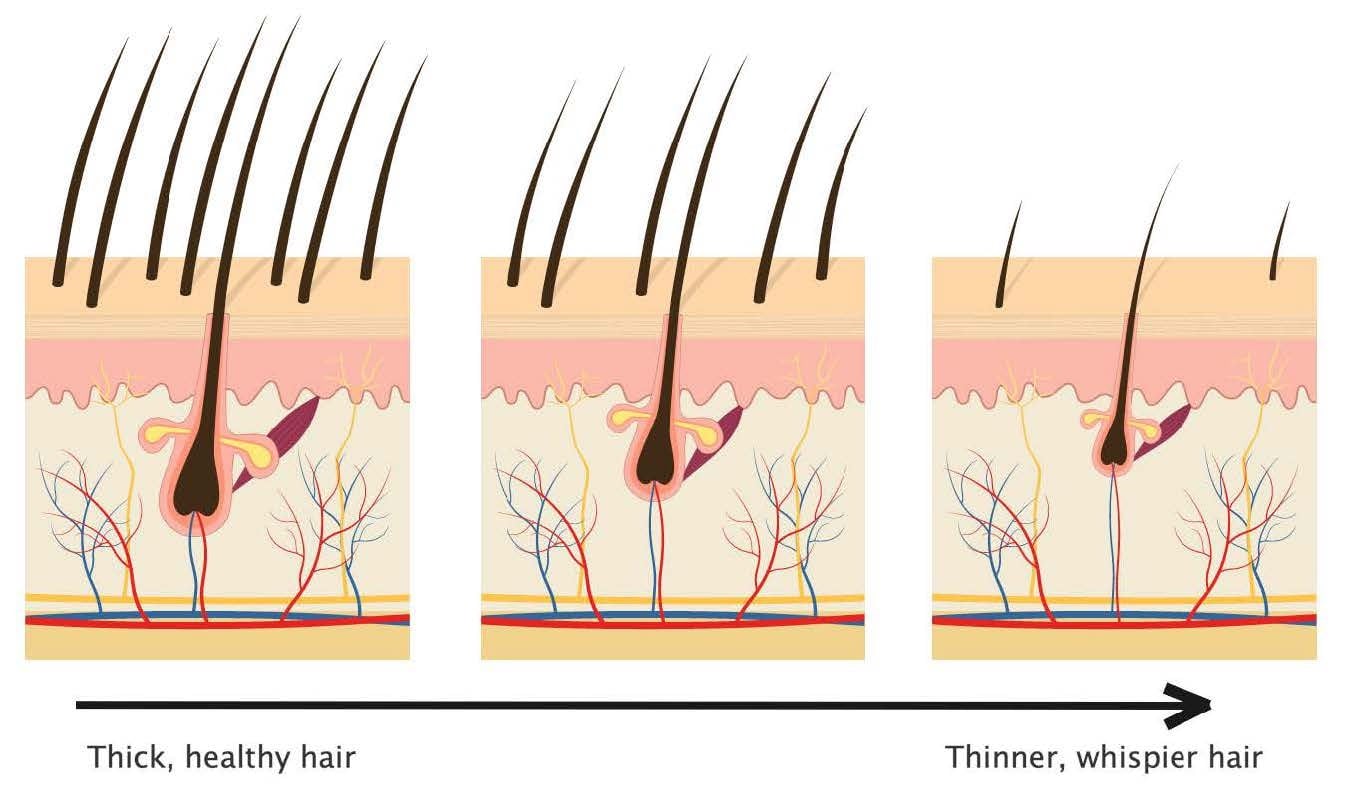
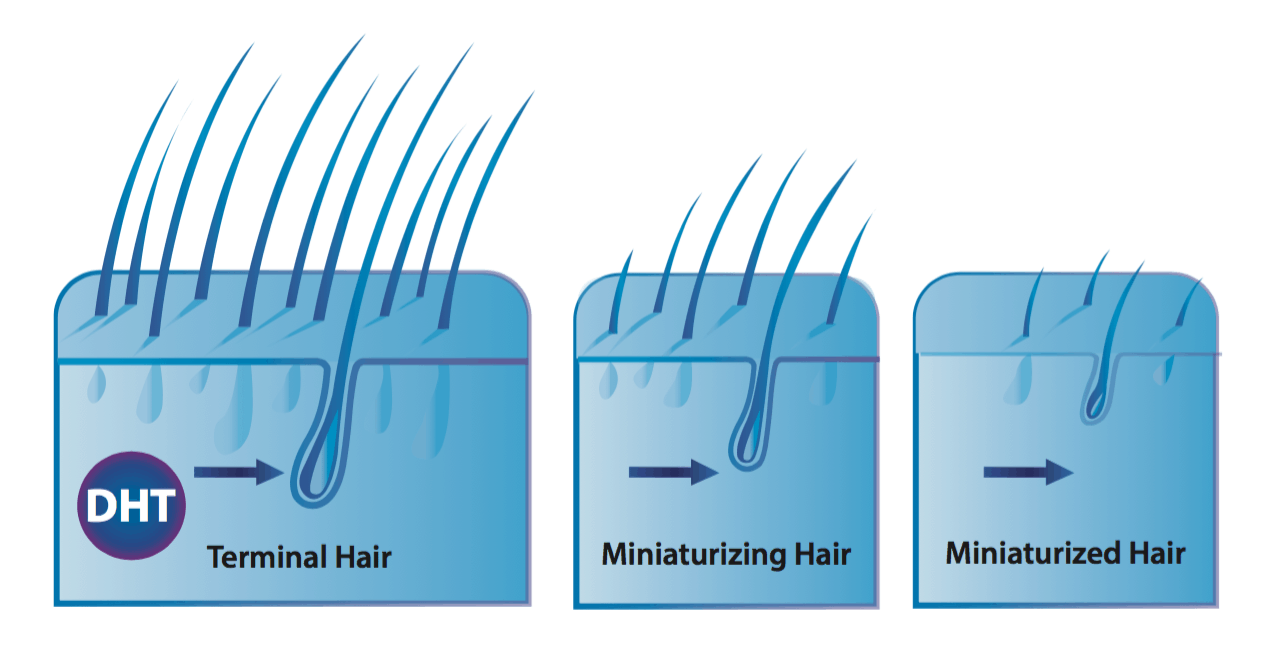
Hair follicle miniaturization
Follicle miniaturization is unique to AGA. It is a process by which AGA-affected hair follicles progressively get smaller until they produce fewer hairs. The hairs that are left may transition into fine, wispy hairs – known as vellus hairs.
Hair follicle miniaturization: a defining characteristic of AGA

While the causes and step-processes of hair follicle miniaturization are still debated, this process is often accompanied by histological (i.e. structural) changes – particularly to the tissues surrounding these hair follicles.
In fact, researchers believe these structural changes help to explain why pattern hair loss is a progressive condition – as they limit miniaturized hair follicles’ capabilities of returning back to full-size. They are:
- Dermal sheath thickening
- Perifollicular fibrosis
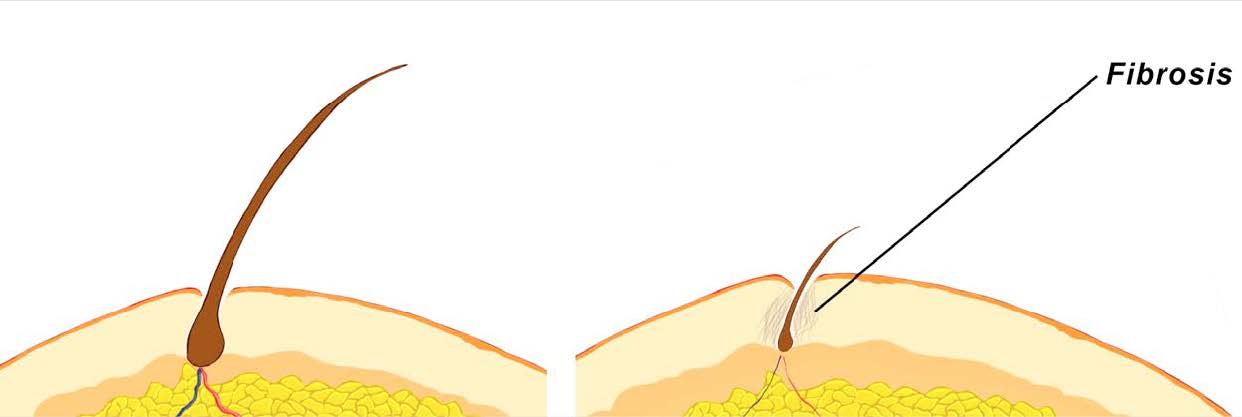
Both of these histological changes can be considered forms of scarring.
Dermal sheath thickening and perifollicular fibrosis
Dermal sheath is a term used to describe the skin tissues that surround our hair follicles. Dermal sheaths are comprised of collagen (i.e., skin), blood vessels, sweat glands, lymphatic networks, and more.
In men and women with AGA, dermal sheaths surrounding miniaturizing hair follicles have thickened. Specifically, their collagen bundles are up to 2.5 times larger – a characteristic of early scar tissue development (Jaworsky et al., 1992).
As dermal sheaths thicken, they widen into the space occupied by hair follicles – thereby impeding their growth space. And as AGA progresses, these dense collagen bundles turn into scar tissue – also known as perifollicular fibrosis.
Together, dermal sheath thickening and perifollicular fibrosis restrict the growth space of surrounding hair follicles, which creates a spacial barrier for miniaturizing hair follicles to recover back to their original size.
Fibrosis (scar tissue) surrounding an AGA-affected hair follicle

Dermal sheath thickening is consistently observed in AGA. However, its advancement to perifollicular fibrosis has been found, according to some studies, in over 71% of AGA sufferers – with moderate levels observed in at least 37% (Whiting et al., 1996).
By progressively restricting the growth space of miniaturizing hair follicles, these histological changes also explain the chronic, progressive nature of AGA – along with why most treatments are limited mainly to stopping the progression of pattern hair loss rather than fully reversing it (English, 2018).
Hair follicle miniaturization is a hallmark of AGA. This is when the size of each hair follicle shrinks and thus starts producing smaller, wispier hairs.
This process is accompanied by dermal sheath thickening around miniaturizing hair follicles. This skin thickening is an early step-process of scarring. In later stages of AGA, this turns to perifollicular fibrosis.
Dermal sheath thickening and perifollicular fibrosis prevent AGA-affected hair follicles from rebounding to their full size. They help to explain the chronic, progressive nature of AGA.
Increased telogen:anagen ratio
Beyond hair follicle miniaturization, the second defining characteristic of AGA is an increase in telogen versus anagen hairs.
Telogen and anagen are terms used to describe specific stages of the hair cycle.
The hair cycle is the natural, ever-repeating cycle of regeneration and degeneration of hair follicles. A hair grows, stops growing, and eventually falls out – at which point a new hair follicle reforms and a new hair starts growing again. In human scalps, these hair cycles last between two to seven years.
The Hair Cycle
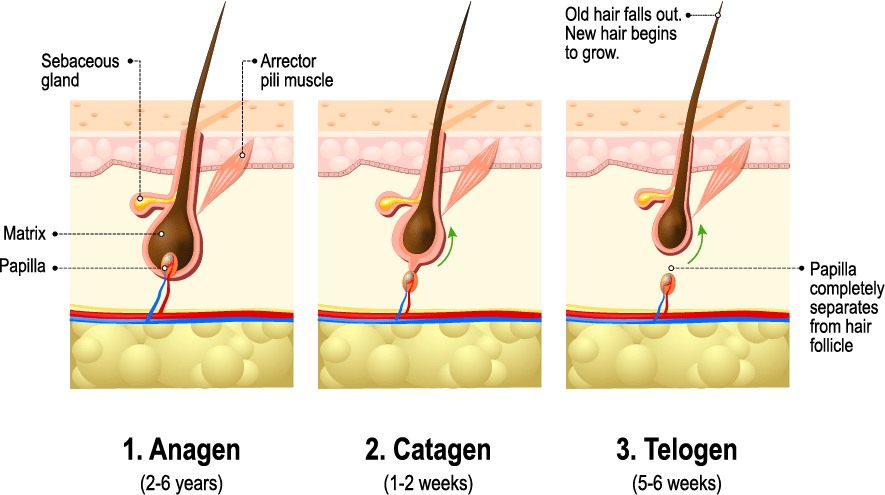

In non-balding scalps, 80-85% of scalp hairs are in their growth (anagen) phase, 1-2% have stopped growing and are in their resting (catagen) phase, and 10-15% have already fallen out and entered their shedding (telogen) phase.
In healthy scalps, this cycle repeats indefinitely. Thus, many hair loss disorders can sometimes be identified by measuring the number of shedding versus growing hairs – and then comparing that ratio to what is seen in normal scalps. If the ratio is high, this suggests abnormal hair loss. If it’s low, this suggests normal, healthy hair.
This is the telogen:anagen ratio.
In normal scalps, there is generally one telogen hair for every 12 anagen hairs, or a 1:12 ratio.
However, in men with AGA, the telogen:anagen ratio can exceed 1:4 (or 25%). In women, this ratio often exceeds 2:5 (or 40%).
Shortened anagen cycling
Moreover, in balding scalps, the anagen phase of a miniaturizing hair is shortened. This means that rather than growing for 2-7 years, these hairs may only grow for a few months (Ho et al., 2019).
This is why so many AGA-affected men and women notice short terminal hairs near their hairline or vertex – hairs that won’t grow more than a few inches. This is the result of a shortened anagen phase and an increased telogen:anagen ratio.
In AGA, balding regions have an elevated ratio of shedding to growing hairs. This is called the telogen:anagen ratio.
While healthy scalps have a telogen:anagen ratio of 10-15%, balding regions will have a ratio of 25-40%, and sometimes higher.
Moreover, growing (anagen) hairs in balding regions often do not grow for more than a few months. This is called a shortened anagen phase. This helps to explain why so many balding men and women see shorter hairs in balding regions that never reach more than an inch or two in length.
What causes AGA?
While the exact step-processes of AGA aren’t yet determined, there is general consensus on two contributing factors: genetics and androgens (i.e., male hormones).
Genetics
AGA has an undisputed genetic component, with one study measuring a 2.5-fold increased risk of developing pattern hair loss in men whose fathers had pattern hair loss, as compared to those whose fathers didn’t (Chumlea et al., 2004). Moreover, women affected by AGA often report having other family members affected by the same condition (Ramos et al., 2015).
However, the exact genes involved in androgenic alopecia have not yet been discovered.
In fact, research suggests that there is no one gene involved in AGA. Rather, pattern hair loss is likely a polygenic disorder, meaning there are many gene variances that are involved in the predisposition of its development.
Scientists are still trying to uncover which polymorphisms may prompt individual susceptibility to AGA. Particular focus is on genes that code for androgen receptors (more on this soon).
Androgen activity
Male hormones (i.e., testosterone) are closely tied to AGA. In the 1940’s, researchers observed that (Hamilton et al., 1942):
- Men castrated before puberty (i.e., before their sex hormones spike) never develop AGA later in life.
- Men with AGA who are castrated (and thereby lose 95% of androgen production) also see a stop in AGA progression.
- Castrated men who are injected with testosterone begin to develop temple recession and/or vertex thinning.
Thirty years later, scientists uncovered the specific male hormone involved in AGA: dihydrotestosterone (DHT).
DHT: the main hormone implicated in pattern baldness
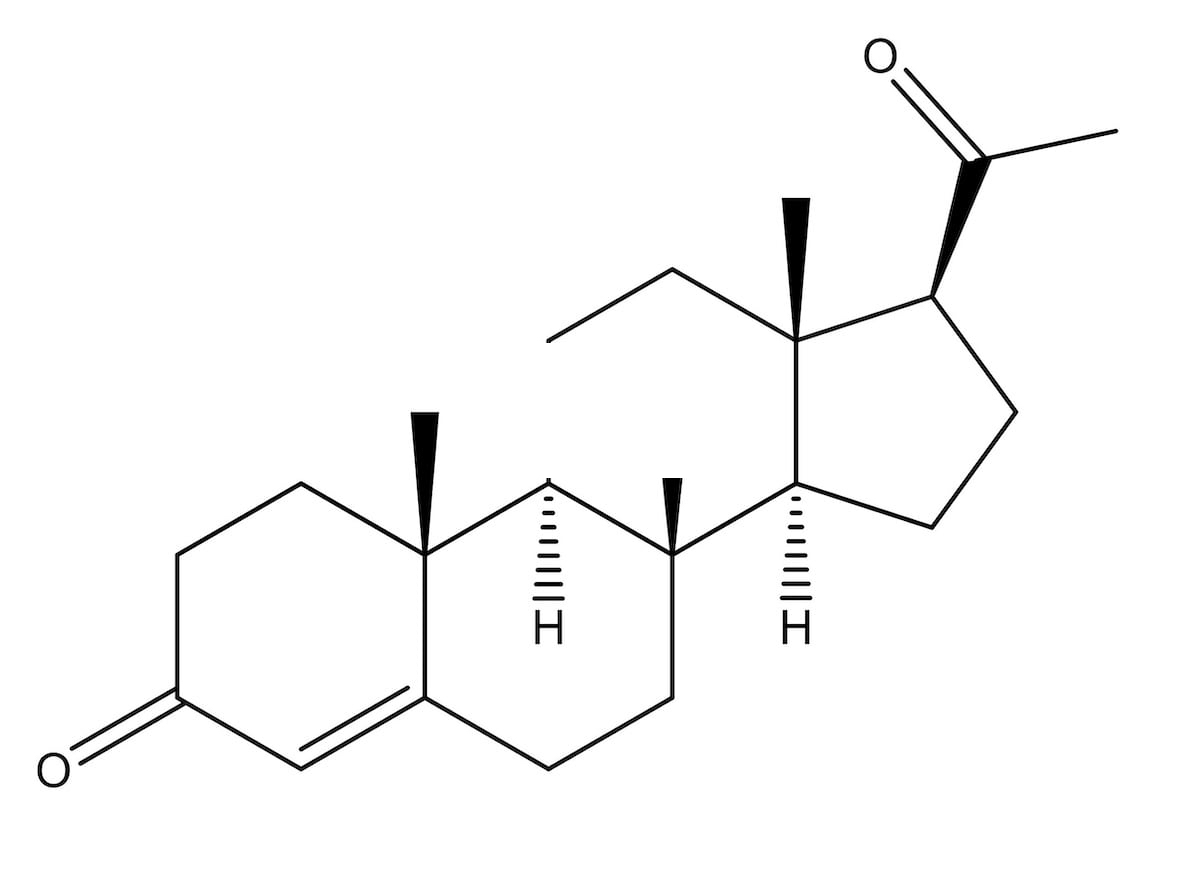

Dihydrotestosterone (DHT)
DHT – a metabolite of testosterone – is causally linked to pattern hair loss (English, 2018).
- Men who cannot make scalp DHT never develop AGA.
- Balding scalp regions have higher amounts of DHT versus non-balding regions.
- Drugs that reduce scalp DHT improve AGA outcomes in 80-90% of men.
DHT drives part of hair follicle miniaturization

However, not all types of DHT are implicated in pattern hair loss. Rather, research is focused more so on one specific type of DHT: DHT made from an enzyme called type II 5-alpha reductase.
Type II 5-alpha reductase
In the body, nearly all DHT is created when unbound testosterone comes into contact with the enzyme 5-alpha reductase. This enzyme binds to free testosterone and then changes testosterone’s structure into DHT.
There are different types of 5-alpha reductase, and most types correspond to a specific tissue or region in which the enzyme expresses (i.e., the skin, brain, prostate, etc.).
When it comes to AGA, the type II 5-alpha reductase is most heavily implicated.
This is because (1) type II 5-alpha reductase is greatly expressed in scalp tissues, and (2) men with a gene mutation who cannot produce type II 5-alpha reductase never go bald (Adachi et al., 1970). Moreover, drugs that inhibit the type II 5-alpha reductase enzyme (i.e., finasteride) help to improve pattern hair loss in men.
Together, these findings implicate DHT and the type II 5-alpha reductase enzyme in AGA. But there’s at least one more androgenic factor involved in the onset and progression of pattern hair loss: that of androgen receptors.
Androgen receptors
When free testosterone comes into contact with type II 5-alpha reductase and converts into DHT, that DHT needs a place to bind to a cell. Once DHT is bound to a cell, this hormone can begin to influence that cell’s functionality.
Androgen receptors are where DHT binds to cells. They’re considered cellular “landing pads” – a place for male hormones to attach, so that they can begin to change cellular behavior.
That means that for (most) scalp DHT to form, we need (1) free testosterone, (2) type II 5-alpha reductase, and (3) an androgen receptor.


This is why androgen receptors are a major focus of AGA research: without androgen receptors, DHT cannot bind to scalp cells and influence their functionality.
“DHT sensitivity”
Type II 5-alpha-DHT has a 5x higher affinity for the androgen receptor than other hormones like testosterone (Trüeb, 2002). This means that in balding scalps, DHT will begin preferentially bind to androgen receptors and exert more effects on cell function, at least compared to other male hormones.
This is why some researchers speak of AGA sufferers developing a “genetic sensitivity” to DHT. In the scalp, the more type II 5-alpha-DHT bound to androgen receptors, the more likely a person is to suffer from pattern hair loss.
Is DHT also involved in female pattern hair loss?
DHT is still involved, but there’s debate over its degree of involvement.
Some studies have demonstrated that both men and women have elevated levels of 5α-reductase in the frontal hair follicles (Orme et al, 1999, Sawaya et al., 1997). This suggests that DHT may play a role in at least some female AGA cases.
At the same time, case studies have demonstrated female pattern hair loss can occur in women who are androgen-deprived, meaning they lack the ability to produce any male hormones at all (Orme et al., 1999).
While some have argued that these instances are just cases of mistaken identity (i.e., not AGA), others have used these findings as starting points to explore other aspects of AGA – and what the DHT-genetic sensitivity argument might not be telling us.
Other potential factors: prostaglandin D2, retinoid receptors, and PPAR pathways
A new wave of AGA research is focusing more so on non-androgenic factors that might influence hair follicle functionality.
In 2014, excitement was generated over prostaglandin D2 and its potential connection to pattern hair loss. Prostaglandin D2 is a fatty acid derivative that, theoretically, can express as a result of androgenic activity (i.e., DHT) (Nieves et al., 2014).
Prostaglandin D2 was once found to be elevated in balding scalp regions and even impede hair lengthening. However, follow-up studies have shown conflicting findings – suggesting that prostaglandin D2 might be less involved in AGA than initially believed (Villarreal-Villarreal et al., 2019).
Interestingly, evidence is accumulating that in addition androgenic activity, retinoid receptors and the PPAR pathway are intimately tied to hair follicle miniaturization – and may even explain why some women develop AGA despite having normal scalp DHT levels (Siu-Yin Ho et al., 2019).
Evidence implicates genes and androgen activity in the onset of AGA.
The hormone DHT is causally linked to AGA. Men who can’t produce DHT never go bald, DHT levels are higher in the scalps of balding men, and drugs that reduce scalp DHT help to stop AGA’s progression.
Scalp DHT is formed when free testosterone comes into contact with the enzyme, type II 5-alpha reductase. This enzyme converts free testosterone into DHT, and then that DHT attaches to an androgen receptor in scalp tissues – thereby influencing cell function.
For these reasons, type II 5-alpha reductase and androgen receptors have been targets of both AGA research and AGA-related drugs. Finasteride is a type II 5-alpha reductase inhibitor; spironolactone and RU58841 are androgen receptor blockers. While these drugs don’t lead to complete AGA reversals, they do seem to improve AGA outcomes.
“DHT sensitivity” is a term used to describe how, in scalp tissues, DHT might begin to preferentially bind to androgen receptors – thereby having up to 5x greater influence over these cells’ behavior than other male hormones like testosterone.
While DHT is causally linked to AGA, it’s still unclear how this hormone causes hair follicle miniaturization. Research in female pattern hair loss brings to question DHT’s involvement in AGA – and suggests that in addition to androgens, other factors must be involved.
What don’t we know about AGA?
Despite AGA’s prevalence and decades of study, there are still a lot of unknowns about this hair loss disorder.
The specific genetics involved
As mentioned, AGA is a polygenic disorder. It has been unequivocally established that male pattern baldness is more likely to occur in men whose fathers suffer from AGA.
However, recent evidence suggests that genetic variances in the gene that encodes for androgen receptors are prevalent among men with male pattern baldness (Ellis et al., 2001). Because the androgen receptor gene is located on the X chromosome, which inherited by men from their mother, this raises questions about whether or not AGA is strictly related to paternal genetics.
Even more confounding is female AGA. Of the genetic components explore in male AGA, none seem to be present in women suffering from pattern hair loss (Ramos et al., 2015).
However, some research has demonstrated is that female pattern hair loss may be associated with polymorphisms in the aromatase gene (aromatase helps synthesize estrogen). Aromatase enzymes help convert testosterone into estrogen. However, it’s still unclear how (or why) these polymorphisms impact both female pattern hair loss – especially as this hair loss disorder has been characterized as “androgenic” since the 1970’s.
From the current body of evidence, it’s almost impossible to distinguish a specific gene or set of genes that is indisputably linked to all cases of AGA.
What causes DHT to increase in balding scalp regions?
If DHT is elevated in balding scalps of most men (and some women), what causes DHT to increase in these regions? This is one question researchers are still trying to answer.
One small-scale study indicates that whole-body DHT may not be the culprit – or that AGA patients don’t seem to have elevated serum DHT versus controls (Urysiak-Czubatka et al., 2014). This gives the impression that elevated DHT in balding scalp tissue is likely a local issue, and not a systemic problem.
So, then, what causes DHT to increase in scalp tissues, specifically? Elevated 5α-reductase activity has been implicated, which explains the ability of 5α-reductase inhibitors like finasteride to halt the progression of AGA (English, 2018). However, finasteride treatment doesn’t fully reverse AGA; it generally just stop its progression.
Findings that AGA is not associated with polymorphisms in the 5α-reductase genes suggest that this enzymatic upregulation is unrelated to genetic predisposition (Ellis et al., 1998). This indicates a possible environmental factor. This assumption is supported by studies showing that in genetically identical twins, one twin can bald faster than his counterpart – despite both twins having the same sets of genes (Nyholt et al., 2003).
Although research is sparse regarding environmental factors influencing 5α-reductase activity, there are some clues.
In women with PCOS, insulin resistance is related to increased 5α-reductase activity, possibly explaining why pattern hair loss is often a feature of the disorder (Wu et al., 2017). Male AGA is also associated with insulin resistance, however, whether this is a result of enhanced enzyme activity is unclear (González-González et al., 2009).
But again, no research team has discovered a definitive answer to this question.
How, exactly, does DHT miniaturize hair follicles?
No one is sure. The closest answers we have (so far) come from DHT and its link to a signaling protein called transforming growth factor beta 1 (TGF-β1).
Follicular miniaturization, as well as dermal sheath thickening and perifollicular fibrosis, are key features of AGA. In vitro studies strongly implicate the role of a growth factor, TGF-β1, in these processes (Yoo et al., 2006).
One of TGF-β1’s primary roles in the body is promoting both wound healing and the deposition of fibrous scar tissue (Pakyari et al., 2013). Thus, elevated TGF-β1 activity is likely involved in AGA follicle miniaturization and may even contribute to the process.
TGF-β1 expression can be triggered by the binding of androgens – like DHT – to androgen receptors, and in a potentially dose-dependent manner (Yoo et al., 2006). TGF-β1 may also enhance androgen activity through an androgen co-activator, Hic-5, that allows androgens like DHT to more effectively influence gene expression, like TGF-β1, within cells (Dabiri et al., 2008).
Essentially, increased androgen activity enhances TGF-β1 expression, and TGF-β1 exacerbates androgen activity. This feedback loop creates a vicious cycle that may underpin AGA progression.
Genetically-determined increased androgen receptor expression in AGA tissue contributes to this upregulated androgen activity, but this genetic component alone isn’t enough to dictate AGA progression (Nyholt et al., 2003).
This begs the question: in AGA, what causes androgen activity to increase?
If activity is dictated by androgen availability in the hair follicle, the rate of DHT conversion by 5α-reductase (which is controlled by the number of 5α-reductase enzymes and their activity), and the involvement of co-activators, then what, beyond genetics, could trigger any one of these factors?
Future research will hopefully begin to answer these questions.
Why is DHT also associated with body and facial hair growth?
DHT is linked to hair loss in AGA scalps. But ironically, this same hormone also enhances facial and body hair growth (Thornton et al.,1993). This occurs in spite of a 3-5 times higher 5α-reductase activity in beard follicles.
Given androgens and their negative impact on hair growth in the scalp, one would assume that elevated 5α-reductase activity would result in beard hair loss, not growth. But, this isn’t true for either case. Why might this be?
Why is there a pattern to androgenic alopecia?
No one is sure. Initially, researchers thought the pattern of AGA was due to different levels of androgen activity in balding regions. However, anecdotes of females with AGA and no androgen activity have raised questions about whether this actually makes sense.
In AGA scalps, there are differences in androgen receptor density and 5α-reductase activity in balding and non-balding regions (Cranwell et al., 2016). This may be explained by in vitro research that proposes individual follicles can “self-regulate”, modulating levels of androgen receptors and 5α-reductase enzymes.
Another question is then: what triggers these follicles to self-regulate? The current state of evidence leaves the answers to these questions purely speculative.
The activation of androgen co-activator, Hic-5, also increases follicle sensitivity to androgens, allowing androgens to more effectively influence hair loss-related gene expression (Tellez-Segura, 2015). Interestingly, stretching forces on follicle cells and oxidative stress have been shown to activate Hic-5 (Kim-Kaneyama et al., 2005, Shibanuma et al., 2003).
Elevated levels of reactive oxygen species (ROS, also known as free radicals) and increased stretching forces in these localized regions may explain increased androgen sensitivity, TGF-β activity, and, thus, the patterning of AGA.
What is not known is what events may trigger increased tension or free radical concentration in these areas. There’s still much to be explored.
The current model of AGA doesn’t fully explain (1) what causes DHT to increase in balding regions, (2) how DHT actually miniaturizes hair follicles, (3) why DHT is associated with scalp hair loss, but body and facial hair growth, and (4) why there is a unique pattern and progression to AGA.
Where is AGA research heading?
As scientists try to answer these questions, they’re beginning to stumble into new (and exciting) areas of AGA research.
Wnt/β-catenin signaling
The hair cycle is a degenerative and regenerative process that requires stem cells. Without the activity of these stem cells, the hair follicle can’t properly regenerate hair or maintain the integrity of the hair follicle itself.
Each compartment of the hair follicle possesses its own stem cell reservoir from which it draws. They are used to repair the epidermis around the hair follicle following injury, maintain the structure of the hair follicle, and help regulate hair follicle cycling (Yang et al., 2010).
Wnt/β-catenin signaling is essential for this process. These pathways help with the differentiation of stem cells in and around the hair follicle.
If this signaling pathway is blocked, hair follicle shaft regeneration is impaired. Blockade of this pathway may also impede the re-entry to the anagen phase from telogen.
In AGA, elevated levels of DHT effectively stimulate androgen-related gene transcription. One of the genes that androgens regulate is dickkopf 1 (DKK-1), a direct inhibitor of the Wnt/β-catenin pathway (Kwack et al., 2008).
When DHT binds to ARs in the dermal papilla cells, it enhances the expression and secretion of DKK-1. It acts in a paracrine fashion, meaning it affects hair components outside of the dermal papilla and may impair stem cell function throughout the hair follicle.
As a result, the hair follicle loses its integrity and may become inactive. However, evidence from animal studies indicates that, thankfully, even long-term DKK-1 inhibition of hair growth may be reversible (Choi et al., 2014).
| Current Model | Alternative Model | |
| How do androgens cause hair loss? | Genetic predispositions | Increased androgen activity and genetic sensitivity work synergistically to inhibit Wnt/β-catenin signaling, preventing hair follicle regeneration and anagen re-entry. |
Galea interaction
The galea aponeurotica is a fibrous connective tissue that extends over the top of the scalp. It is attached to and connects all the muscles surrounding it.
The galea is a key component of the scalp tension theory of AGA. Interestingly, the “pattern” observed in AGA directly correlates to the highest points of tension in the GA (English, 2018).
Hair follicles are fashioned within both the dermis and fatty tissue beneath the skin, which is directly fused with the GA. Tension in the galea may be transmitted to these tissues and the follicles that are housed within.
Stretch-induced mechanical tension can enhance free radical production and trigger chronic inflammation in various types of tissues, potentially including the GA and the tissues that surround it. Cellular mechanical tension, free radicals, and inflammation can all enhance androgen activity and the fibrosis-mediating TGF-β1 (English, 2018).
This may, in part, explain the origin of follicle miniaturization in AGA.
Other research groups have developed similar hypotheses, arguing that fibrosis may not only drive hair follicle miniaturization, but that this process is potentially mediated by interactions between the galea and cells that transition adipose (fat) tissue into scar tissue (Kruglikov et al., 2017).
| Current Model | Alternative Model | |
| What causes increased androgen activity in AGA follicles? | Genetic predisposition to androgen sensitivity. | Mechanical stretch, free radical production, and chronic inflammation as a result of galea tension enhance androgen activity. |
| Why is there increased androgen activity in localized regions? | Genetics don’t yet offer any direct explanation. | The points of highest tension within the galea directly correlate to the pattern seen in AGA. The mechanical stretch, free radical production, and chronic inflammation may all upregulate androgen activity. |
| Why does DHT cause hair loss on the scalp but not in facial or body hair? | Genetics don’t yet offer any explanation. | Mechanical stretch, free radical production, and chronic inflammation as a result of GA tension trigger TGF-β1 expression that leads to follicle miniaturization. |
Olfactory receptors
An interesting finding of one 2018 study was the identification of OR2AT4 receptors in human hair follicles. OR2AT4 is an olfactory or smell receptor that is activated by certain scents (Chéret, et al. 2018).
In this study, researchers demonstrated that activation of this receptor by a synthetic sandalwood scent prolonged the anagen phase of the hair cycle. In fact, their findings suggest that activation of OR2AT4 may actually be indispensable for maintaining anagen.
While OR2AT4 is primarily considered an olfactory receptor activated by scent, this study also suggests that it may be activated by other compounds such as scalp microflora (bacteria) metabolites and short-chain fatty acids.
| Current Model | Alternative Model |
| Inhibition of androgen activity is the best way to treat AGA. | Many different cellular receptors in the hair follicle, like olfactory receptors, may help counteract the effects of androgens in the scalp. |
In addition to PPAR pathways and retinoid receptors – Wnt-β catenin signaling, galea interactions, and olfactory receptors may help explain much of the unexplained phenomena in AGA pathology.
What are the treatment targets for AGA?
Most AGA treatments target to (1) decrease the telogen:anagen ratio, and (2) stop the progression of hair follicle miniaturization.
There are many ideas as to how to do this, but the most successful FDA-approved approach (so far) seems to be finasteride (Propecia®), a type II 5-alpha reductase inhibitor.
1mg daily of oral finasteride reduces scalp DHT levels by 50-70%. Subsequently, it stops AGA progression in 80-90% of men and leads to a 10% increase in hair count over two years, along with some additional hair thickening (English, 2018).
However, prolonged finasteride use is sometimes associated with sexual side effects. And, when considering all of the evidence that not just androgens are involved in AGA, it’s likely that finasteride is not a complete solution.
As such, here’s a list of current treatment targets in AGA research. Many of these targets overlap with one another.
- Type II 5-alpha reductase
- Androgen receptors
- Reactive oxygen species
- Wnt-β catenin pathways
- DKK-1
- TGF-β1
- Prostaglandin analogues
- Olfactory receptors
- PPAR pathways
- Retinoid receptors
- Potassium ion channels
- Mechanical tension
- Adipose-myofibroblast transitions
- Scalp microflora
In the years to come, this list will undoubtedly grow as researchers elucidate more of the molecular mechanisms behind AGA.
The bottom line
AGA can affect men and women, in both similar and completely different patterns.
AGA is characterized by (1) an increased telogen:anagen ratio, and (2) hair follicle miniaturization. Dermal sheath thickening and perifollicular fibrosis are also present in balding regions, and may partly explain the chronic, progressive nature of AGA.
Both genetics and androgens are established as causally linked to AGA. Androgen receptor density, type II 5α-reductase activity, and DHT are elevated in balding scalps.
The DHT “genetic sensitivity” argument for AGA is incomplete, and for several reasons. (1) Elevated whole-body DHT and polymorphisms in the 5α-reductase gene are not associated with AGA. (2) Female pattern hair loss has been observed in women who cannot produce androgens. (3) More recent research now implicates non-androgenic factors like retinoid receptors and PPAR pathways in the onset of hair follicle miniaturization from AGA.
The current model of AGA doesn’t fully explain (1) what causes DHT to increase in balding regions, (2) how DHT actually miniaturizes hair follicles, (3) why DHT is associated with scalp hair loss, but body and facial hair growth, and (4) why there is a unique pattern and progression to AGA.
Wnt-β catenin signaling, galea interactions, and olfactory receptors are a few new research areas in AGA that might help explain part of the unexplained phenomenon in AGA.
(1) Wnt/β-catenin signaling regulates the stem cell activity in hair follicles needed for follicle regeneration and anagen re-entry. Androgens stimulate DKK-1 secretion by dermal papilla cells which inhibits Wnt/β-catenin signaling and impairs stem cell activity.
(2) Mechanical tension in the galea, which directly corresponds to the patterning in AGA, may play some sort of regulatory role in the inflammation, free radical production, and TGF-β1 expression that seems to mediate scarring and hair follicle miniaturization. However, research here is limited.
(3) Activation of one olfactory receptor by short-chain fatty acids, scalp microflora metabolites, and specific scents may prolong the anagen phase, providing one possible avenue for counteracting AGA progression outside of the traditional anti-androgen treatment approach.
AGA researchers are now turning focus toward non-androgenic factors – PPAR pathways, retinoid receptors, and more – to explain the unexplainable in AGA pathology. If research keeps heading in this direction, newer treatment options will likely better target AGA’s step-processes, resulting in better hair recovery and with a greatly reduced potential for side effects.


Rob English is a researcher, medical editor, and the founder of perfecthairhealth.com. He acts as a peer reviewer for scholarly journals and has published five peer-reviewed papers on androgenic alopecia. He writes regularly about the science behind hair loss (and hair growth). Feel free to browse his long-form articles and publications throughout this site.